
Private: Appendices
17 Osteology
Jason M. Organ, Ph.D., Indiana University School of Medicine
Jessica N. Byram, Ph.D., Indiana University School of Medicine
Learning Objectives
- Identify anatomical position and anatomical planes, and use directional terms to describe relative positions of bones.
- Describe the gross structure and microstructure of bone as it relates to bone function.
- Describe types of bone formation and remodeling, and identify (by name) all of the bones of the human skeleton.
- Distinguish major bony features of the human skeleton like muscle attachment sites and passages for nerves and/or arteries and veins.
- Identify the bony features relevant to estimating age, sex, and ancestry in forensic and bioarchaeological contexts.
- Compare human and chimpanzee skeletal anatomy.
Anthropology is the study of people, and the skeleton is the framework of the person. So while all subdisciplines of anthropology study human behavior (culture, language, etc.) either presently or in the past, biological anthropology is the only subdiscipline that studies the human body specifically. And the fundamental core of the human (or any vertebrate) body is the skeleton. Osteology, or the study of bones, is central to biological anthropology because a solid foundation in osteology makes it possible to understand all sorts of aspects of how people have lived and evolved. For example, bones from archaeological sites can be used to understand what animals people ate; fossilized bones can tell us how human and other primate locomotion has changed through time; and bones can give us clues to how modern and ancient humans died, whether by natural causes or in the context of forensic investigations. This appendix will introduce the reader to the basics of anatomical terminology and will then describe the different regions and bones of the skeleton with an emphasis on those structures that have evolved dramatically in humans compared to our closest living relatives: chimpanzees. It will also highlight some (but not all) of the features of the skeleton (e.g., bumps and grooves on bones) used in a forensic context to estimate the age and sex of recovered remains.
ANATOMICAL TERMINOLOGY
Scientists have adopted standardized terminology to describe the position of the body as well as the location and movements of different body parts relative to one another. The terminology used throughout this appendix is consistent with the most recent edition of Terminologia Anatomica: International Anatomical Terminology (Federative Committee on Anatomical Terminology 1998). Below you will find descriptions of standard anatomical position, directional terms, anatomical planes, and skeletal movements (i.e., movements possible where two bones articulate through a joint).
When a body is in anatomical position, it is situated as if the individual is standing upright; with head, eyes, and feet pointing forward (anteriorly, see below); and with arms at the side and palms facing forward. In anatomical position, the bones of the forearm are not crossed (Figure A.1).

With the body in anatomical position, the position of specific organs (e.g., bones) can be described as situated within specific anatomical planes (Figure A.2). These imaginary planes bisect the body into equal or subequal halves, depending on which plane is described. Coronal (frontal) planes divide the body vertically into anterior (front) and posterior (back) halves. Transverse planes divide the body horizontally into superior (upper) and inferior (lower) halves. Sagittal planes divide the body vertically into left and right halves. The plane that divides the body vertically into equal left and right halves is called the midsagittal plane. The midsagittal plane is also called the median plane because it is in the midline of the body. If the left and right halves of the body are divided unequally (i.e., the right “half” is larger than the left “half” or vice versa), we call that dividing plane a parasagittal plane. There are potentially an infinite number of parasagittal planes that can be drawn to divide the body into unequal right and left “halves.”

Directional Terms
When describing the position of one body part (in this case, a bone) relative to another, scientists use precisely defined directional terms. Each of the directional terms described below refers to the body in anatomical position. This is an important point because once the position of one bone is established relative to another, that directional term is the same, regardless of whether the body remains in anatomical position (e.g., the skull is always superior to the vertebrae, even if the individual is lying down).
A bone or skeletal feature that is anterior (or ventral) is located toward the front of the body and a bone that is posterior (or dorsal) is located toward the back of the body (Figure A.3). For example, the sternum (breastbone) is anterior to the vertebral column (“backbone”). A medial bone is located closer to the midline (midsagittal plane) than a bone that is lateral, or located further from the midline. For example, the thumb is lateral to the index finger. A structure that is proximal is closer to the trunk of the body (usually referring to limb bones) than a distal structure, which is further from the trunk of the body. For example, the femur (thigh bone) is proximal to the tibia (leg bone). Finally, structures that are superior (or cranial) are located closer to the head than structures that are inferior (or caudal). For example, the rib cage is superior to the pelvis, and the foot is inferior to the knee. Typically, the terms “cranial” and “caudal” are used in reference to the non-human, quadrupedal skeleton, whereas “superior” and “inferior” are used in reference to the human skeleton.

SKELETAL FORM AND FUNCTION
Structure and Material of Bone

Bone is a composite of organic (collagen) and inorganic (mineral, e.g., hydroxyapatite, a calcium phosphate salt) materials with incredible strength in compression so it can support the body under the force of gravity. When bone is mature (fully mineralized as opposed to juvenile and undermineralized), it comprises an outer dense region of bone called cortical (or compact) bone and an inner spongy region of bone called cancellous (or trabecular) bone (Figure A.4). However, the interfaces between the organic and inorganic materials, as well as the cortical and cancellous regions, are subject to changing stresses. Each time we move our muscles, our bones are subjected to a combination of bending, twisting, compression, and tension. This results in the formation of microscopic cracks that weaken the bone and may result in complete bone fracture. Bone cells called osteocytes have special properties that allow them to sense when these microcracks form. Osteocytes then signal osteoclast cells to remove the cracked bone and osteoblast cells to lay down new bone—a process known as skeletal remodeling. Osteogenic cells are stem cells that are able to differentiate into osteoblasts (Figure A.5).

Bone Shape
Different bones have different shapes that largely relate to their specific function within the skeletal system. Additionally, the ratio of cortical to cancellous bone, and which muscles are attached to the bone and how, affect the shape of the whole bone. Generally there are five recognized bone shapes: long bones, short bones, flat bones, sesamoid bones, and irregular bones. Long bones are longer than they are wide and consist of three sections: diaphysis, epiphysis, and metaphysis (Figure A.4). The diaphysis of a long bone is simply the shaft of the bone, and it comprises mostly cortical bone with a thin veneer of internal cancellous bone lining a medullary cavity. At both the proximal and distal ends of every long bone, there is an epiphysis, which consists of a thin shell of cortical bone surrounding a high concentration of cancellous bone. The epiphysis is usually coated with hyaline (or articular) cartilage to facilitate joint articulation with other bones. The junction between diaphysis and epiphysis is the metaphysis, which has a more equal ratio of cortical to cancellous bone. Examples of long bones are the humerus, the femur, and the metacarpals and metatarsals.
The other bone shapes are simpler. Short bones are defined as being equal in length and width, and they possess a mix of cortical and cancellous bone (Figure A.6). They are usually involved in forming movable joints with adjacent bones and therefore often have surfaces covered with hyaline cartilage. Examples of short bones are the carpals of the wrist and the tarsals of the ankle.
Flat bones, as their name suggests, are flat and consist of two layers of thick cortical bone with an intermediate layer of cancellous bone called a diploe. Examples of flat bones are most of the bones of the skull, such as the frontal and parietal bones, as well as all parts of the sternum (Figure A.6). Sometimes bones develop within the tendon of a muscle in order to reduce friction on the joint surface and to increase leverage of the muscle to move a joint. These types of bones are called sesamoid bones, and these include the patella (or knee cap) and the pisiform (a bone of the wrist).
Irregular bones are bones that don’t fit into any of the other four categories. The shapes of these bones are often more complex than the others, and examples include the vertebrae and certain bones of the skull, like the ethmoid and sphenoid bones (Figure A.6).

Bone Formation
Bone develops via one of two mechanisms: intramembranous or endochondral bone formation. Intramembranous bone formation occurs when connective tissue mesenchymal (stem) cells aggregate and differentiate into osteoblasts, which then begin to synthesize new bone along the aggregated connective tissue cells (Figure A.7). Intramembranous bone formation is the mechanism by which most bones of the skull develop as well as the clavicle (collar bone). When osteoblasts develop from an intermediate cartilage “model” that is then replaced by bone, instead of developing directly from the mesenchymal cells, the mechanism is described as endochondral bone formation (Figure A.8). Endochondral bone formation is the mechanism by which most bones of the skeleton develop (Burr and Organ 2017).


Bone Function
Bone performs both metabolic and mechanical functions for the body. On the metabolic side, bone is required to maintain mineral (i.e., calcium) homeostasis and for the production of red and white blood cells (Figure A.9), which develop in the cavity and the cancellous region of the metaphysis and epiphysis. But it is undeniable that the mechanical functions of bone are primary because bone is critically responsible for protecting internal organs, providing support against the force gravity, and serving as a network of rigid levers for muscles to act upon during movement.

HUMAN SKELETAL SYSTEM
The skeletal system is divided into two regions: axial and appendicular (Figure A.10). The axial skeleton consists of the skull, vertebral column, and the thoracic cage formed by the ribs and sternum (breastbone). The appendicular skeleton comprises the pectoral girdle, the pelvic girdle, and all the bones of the upper and lower limbs (White and Folkens 2000).
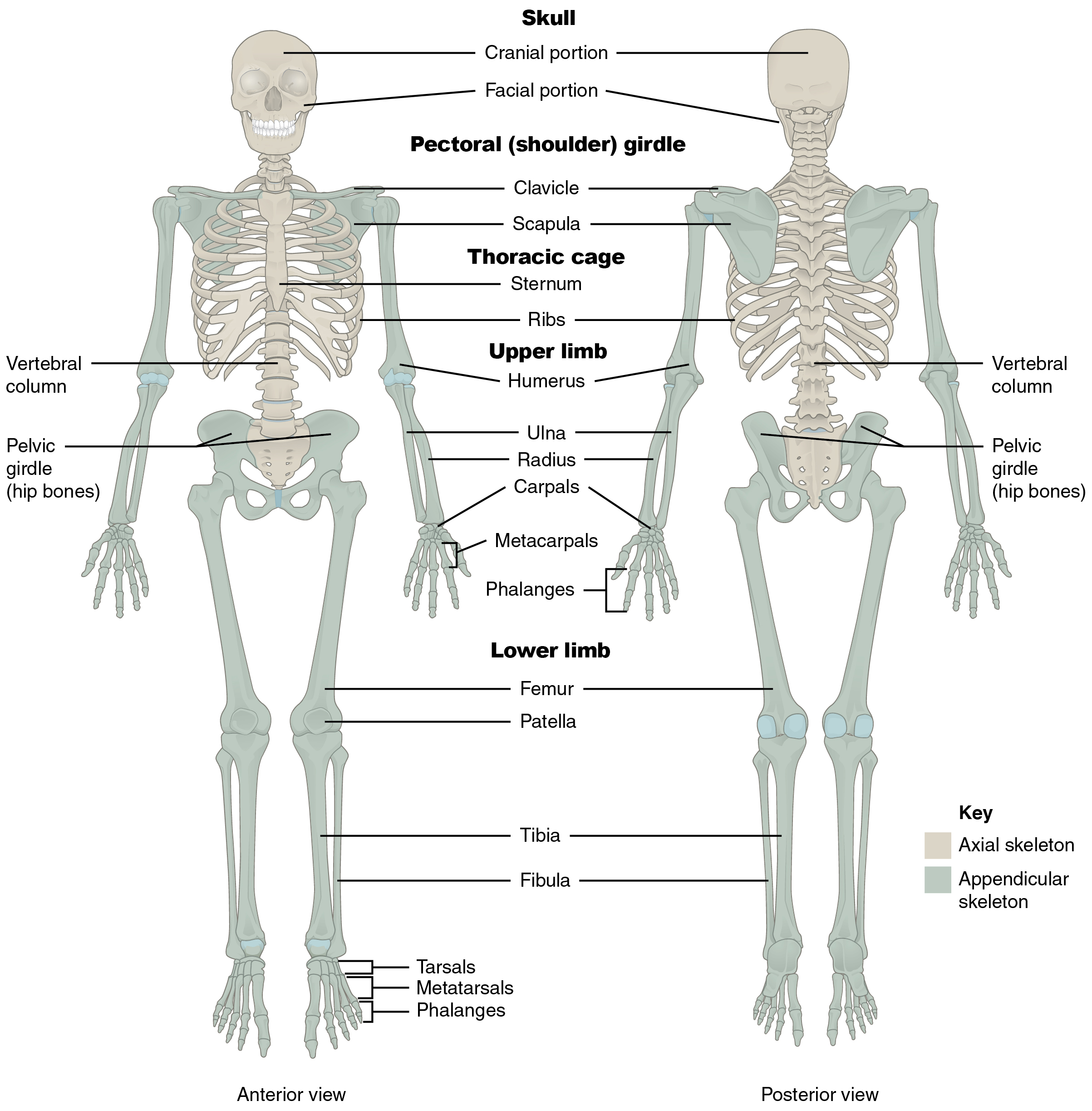
Axial Skeleton
Skull
The skull comprises numerous bones (some paired and others that are unpaired) and is divided into two major portions: the mandible (or lower jaw) and the cranium (the remainder of the skull). The cranium is further subdivided into the neurocranium (or cranial vault), which houses the brain, and the viscerocranium (or facial skeleton; Figure A.11). Where two bones of the cranium come together, they form articulations called cranial sutures, which fuse (or close) with increasing age and can be used as a broad estimate of age at death. Degree of suture closure is scored at several anatomical landmarks and compiled to produce an age estimate. The remainder of this section lists the bones of the skull by region and details some of the landmarks examined in forensic contexts.

Bones and Some Features of the Neurocranium
- Frontal: an unpaired bone consisting of two parts: a superior, vertically oriented portion called the squama and an inferior, horizontally oriented portion that forms the roof of the orbit (eye socket; Figures A.12 and A.13).
- The coronal suture is the articulation between the frontal bone and the two parietal bones posterior and lateral to the frontal.
- The frontal bone develops initially as two separate bones that fuse together during growth. Occasionally this fusion is incomplete, resulting in a metopic suture that persists between the two halves (left and right) of the frontal bone (White and Folkens 2005).
- The glabella is a bony projection between the brow ridges. The glabella in females tends to be flat while rounded and protruding in males.
- The supraorbital margin is the upper edge of the orbit. The thickness of the edge can be used as an indicator of sex. A thin, sharp border is indicative of a female while a blunt, thick border suggests a male.


- Parietal: Paired bones that form the majority of the roof and sides of the neurocranium (Figures A.12 and A.13).
- The sagittal suture is the articulation between the right and left parietal bones. It extends from the coronal suture anteriorly to the lambdoidal suture, which separates the parietal bones from the occipital bone posteriorly.
- Each parietal bone is marked by two temporal lines (superior and inferior), which are anterior-posterior arching lines that serve as attachment sites for a major chewing muscle (temporalis) and its associated connective tissue.
- Temporal: Paired bones on the lateral side of the neurocranium that are divided into two portions: squamous (or flat) portion that forms the lateral side of the neurocranium and the petrous (or rock-like) portion that houses the special sense organs of the ear for hearing and balance as well as the three tiny bones of the middle ear: incus, malleus, and stapes (Figures A.13, A.14, and A.15).
- The squamosal suture is the articulation between the squamous portion of the temporal bone and the inferior border of the parietal bone.
- The mastoid process is a prominent attachment site for several muscles including the large sternocleidomastoid muscle. As such, it is often used to estimate sex in that males tend to have longer and wider mastoid processes compared to females (Bass 2005).
- The styloid process is a thin, pointed, inferior projection of the temporal bone that serves as an attachment site for several muscles and a ligament of the throat.
- The zygomatic process of the temporal is a thin, arch-like process that originates from the squamous portion of the temporal bone. The zygomatic process articulates with the temporal process of the zygomatic bone to form the zygomatic arch (or cheekbone).
- The temporal fossa is the depression in the temporal bone where the mandibular condyle (see below, under mandible) articulates to form the temporomandibular (or jaw) joint.


- Occipital: Unpaired bone that forms the posterior and inferior portions of the neurocranium (Figures A.13 and A.15).
- The lambdoidal suture is the articulation between the occipital bone and the two parietal bones. It resembles the shape of the Greek letter lambda.
- The external occipital protuberance (EOP) is a robust attachment of the nuchal ligament. When viewed laterally, males tend to have a discernible projection or hook, whereas in females, the occipital is typically smooth.
- The nuchal lines are parallel ridges that meet on the midline at the EOP. These prominent muscle attachment sites tend to be more robust and projecting in males.
- The occipital bone contains a large circular opening called the foramen magnum, which provides a space for passage of the brainstem/spinal cord from the neurocranium into the vertebral canal of the spine.
- Sphenoid: Unpaired, butterfly-shaped bone that forms the central portion of the bottom of the neurocranium. The sphenoid is divided into several regions, including the body, greater wings, lesser wings, and pterygoid processes (with pterygoid plates; see Figures A.15 and A.16). This bone is critical to supporting the brain and several nerves and blood vessels supplying this region.
- Pterygoid plates are flat projections of the pterygoid processes that serve as attachment sites for chewing muscles and muscles of the throat.

- Ethmoid: Unpaired bone consisting of a median vertical plate that forms part of the bony nasal septum and a horizontal plate (cribriform plate) with many small foramina (holes) that transmit olfactory nerves (special sense of smell; Figure A.17).

Bones of the Viscerocranium
- Maxilla: Paired bones that form the upper jaw, support the upper teeth, and form the inferior margin of the cheek (Figures A.12, A.13, A.15, and A.18).
- The nasal spine is a thin projection on the midline at the inferior border of the nasal aperture. Length of the nasal spine has been used as a subjective trait to determine ancestry, with those exhibiting long nasal spines to be more likely of European ancestry.
- The zygomatic process of the maxilla is the portion of the bone that articulates with the zygomatic bone to form the anterior portion of the zygomatic arch.

- Nasal: Small, paired, flat, rectangular bones that form the bridge of the nose (Figure A.19).
- Nasal aperture is the anterior opening into the nasal cavity. As a forensic trait, it is described as low and wide in those of African ancestry and tall and narrow in those of European ancestry.
- Zygomatic: Paired bones that form the anterolateral portion of the cheekbone and contribute to the lateral and inferior wall of the orbit (Figure A.19).
- The temporal process of the zygomatic is the portion of the bone that articulates with the temporal bone to form the anterior portion of the zygomatic arch.
- Palatine: Paired L-shaped bones that form the posterior portion of the roof of the mouth, floor of the orbit, and the floor and lateral walls of the nasal cavity (Figures A.15 and A.19).
- Lacrimal: Small, flat, paired bones that form the anterior portion of the medial wall of the orbit (Figure A.19).


- Vomer: Unpaired thin bone that forms the inferior portion of the bony nasal septum. It articulates with the ethmoid superiorly (Figure A.20).
- Inferior nasal concha: Paired bones that project and curl like a scroll from the lateral wall of the nasal cavity (Figure A.21).

- Hyoid: Unpaired U-shaped bone that sits in the neck inferior to the mandible. The hyoid is the only bone of the skeleton that does not articulate with another bone. Instead, it is encased in a sling of muscles that move the larynx (voice box), pharynx, and tongue (Figure A.22).

- Mandible: Unpaired bone with a horizontal (and anteriorly arched) body and a vertical ramus that articulates with the temporal fossa to form the temporomandibular (jaw) joint. The body of the mandible houses the lower teeth (Figure A.13 and A.23).
- The mental protuberance (eminence) is the most anteriorly projecting point on the mandible—the so-called “chin.” A prominent projection is likely to indicate a male, while a smooth mental region likely indicates a female.
- The ramus of the mandible projects superiorly from the body of the mandible and ascends to one of two features on the superior aspect: coronoid process or mandibular condyle.
- The coronoid process is a bony projection off the anterior and superior aspect of the mandibular ramus. The inferior attachment of the temporalis muscle (a chewing muscle) attaches here.
- The mandibular condyle, a rounded projection off the posterior and superior aspect of the mandibular ramus, articulates with the temporal fossa of the temporal bone at the temporomandibular (TMJ) joint.
- The gonial (mandibular) angle is the rounded posteroinferior border of the mandible. It tends to be smooth in females with a more obtuse angle but is laterally flared in males and closer to a right angle in shape.

- Teeth: Adults normally have 32 teeth, distributed among four quadrants of the mouth (upper left, upper right, lower left, lower right). In each quadrant, there are eight teeth: two incisors (central and lateral), one canine, two premolars, and three molars. Each of these types of teeth has a different shape that reflects its function during chewing:
- Incisors are flat and shovel shaped and are used to bite into a food item.
- Canines are conical, with a single pointed cusp used to puncture a food item.
- Premolars have two rounded cusps and are used to grind and mash a food item.
- Molars have five flatter cusps and are used to grind food prior to swallowing.
The teeth have their own set of directional terms that help differentiate the different parts of the tooth. For example, the anterior portion of the tooth is called mesial, while the posterior portion of the tooth is called distal. In the case of teeth in the front of the mouth, mesial refers to the aspect toward the midline of the body; distal refers to the aspect away from the midline. Similarly, the side of the tooth facing the lips is called the buccal surface and the side facing the tongue is called the lingual surface. Finally, we can talk about the occlusal surface of the tooth, which is the surface that comes in contact with food or the teeth from the other jaw when the jaw is closed. Sometimes the occlusal surface of the incisors is called the incisal surface.
Vertebral Column
The adult vertebral column consists of 32–33 individual vertebrae, divided into five regions: cervical, thoracic, lumbar, sacral, and coccygeal. This section will review the features of a general vertebra and then will describe the unique features of vertebrae in each of the five regions.
General Structure of a Vertebra
A typical vertebra consists of an anteriorly situated body—the main weight-bearing element of the vertebra—and a posteriorly projecting vertebral arch (Figure A.24). The vertebral arch consists of the paired pedicles and paired laminae. The pedicle connects the transverse process (a laterally projecting process that serves as an attachment site for muscles and ligaments) to the vertebral body; the lamina connects the spinous process (a posteriorly projecting process that serves as an attachment site for muscles and ligaments) to the transverse process. Projecting inferiorly off the vertebral arch is the inferior articular process, and projecting superiorly off the vertebral arch is the superior articular process. Between the vertebral body anteriorly and the vertebral arch posteriorly is an open space called the vertebral foramen.
Adjacent vertebrae articulate with one another through two major types of joints: intervertebral disc joints between adjacent vertebral bodies and zygapophyseal (facet) joints between the inferior articular process of one vertebra and the superior articular process of the vertebra immediately inferior to it. When all vertebrae are articulated into a column, the adjacent vertebral foramina form the vertebral canal, through which the spinal cord travels from the foramen magnum of the occipital bone to approximately the level of the second lumbar vertebra. At the level of each vertebra, the spinal cord gives us a pair (left and right) of spinal nerves that exit between adjacent vertebrae through the intervertebral foramen formed by adjacent vertebral arches. Even though the spinal cord ends in the lumbar region, the spinal nerves emanating from the spinal cord continue all the way to the sacrum (sometimes to the coccyx), culminating in a total of 30–31 pairs of spinal nerves.

Regional Differences in Vertebral Shape
In the cervical region of the vertebral column, there are seven vertebrae (named C1–C7 from superior to inferior; Figure A.25). The first two cervical vertebrae are unique from each other and all other cervical vertebrae, and they get special names: atlas (C1) and axis (C2). The atlas lacks a vertebral body (having only two large articular facets for articulation with the occipital bone of the skull: the atlanto-occipital joint for nodding the head) and does not have a spinous process. The axis is notable for the superiorly projecting dens (or odontoid process), which articulates with the atlas to create the atlanto-axial joint for head rotation. Otherwise, a typical cervical vertebra has a small vertebral body, a bifid (split) spinous process, a transverse process with a transverse foramen on it for passage of the vertebral artery and vein, and a triangular-shaped vertebral foramen.

The vertebrae in the other regions of the spinal column are less variable in shape than the cervical region vertebrae. There are 12 thoracic region vertebrae (T1–T12), and they can be easily distinguished from the vertebrae in other regions because they have articular facets on their vertebral bodies for articulation with the head of a rib, as well as articular facets on the transverse process for articulation with the rib tubercle (Figure A.26). In particular, the vertebral bodies of T2–T9 have two pairs of articular facets called demifacets (superior and inferior), for articulation with multiple ribs; T1 and T10–T12 have single facets for articulation with a single rib. All five lumbar region vertebrae (L1–L5) are distinguished by their large vertebral body and rounded spinous process (Figure A.27). Finally, there is the sacrum, which is a bone of the pelvis that forms from the fusion of all five sacral region vertebrae (S1–S5), and there is the coccyx, which comprises three to four fused coccygeal region vertebrae that form the tailbone (Figure A.28). All great apes (including humans) lack an external tail; the coccygeal vertebrae are homologs of the external tail vertebrae in other primates and mammals (Organ 2017). Homologs are anatomical features that have the same evolutionary origin but do not necessarily have identical structure or function (i.e., the wings of bats and the arms of humans are homologous).



Curvatures of the Vertebral Column
The adult spine is curved in the midsagittal plane in four regions of the vertebral column (cervical, thoracic, lumbar, and sacral; Figure A.29). During the fetal period of development, the vertebral column develops an anteriorly concave curvature called a kyphosis. But during the postnatal period, when an infant learns to hold its head up and then again when it learns to walk, it develops secondary curvatures called lordoses (singular: lordosis) that are posteriorly concave in the cervical and lumbar vertebral regions, while the kyphoses remain in the thoracic and sacral regions. The end result is an S-shaped curvature to our spine that enables us to keep our head and torso above our center of mass (near our pelvis) while walking around on two legs.

Thoracic Cage
The thoracic cage is formed from the sternum and the 12 ribs and their cartilages (costal cartilages), and the 12 thoracic vertebrae with which the ribs articulate (Figure A.30). The sternum comprises the manubrium (superior portion), the body of the sternum, and the xiphoid process. Each rib has a head and neck (with rib tubercle) at the vertebral end of the rib as well as a flattened shaft that extends to articulate with the sternum. All ribs articulate with the vertebral column at two points: the transverse process facet (rib tubercle) and vertebral body articular facet (head of rib). But articulations between the ribs and the sternum vary, where some ribs (1–7, the “true ribs”) attach directly to the sternum via their costal cartilages, other ribs (8–10, the “false ribs”) attach indirectly to the sternum via the costal cartilage of the rib above, and some ribs (11–12, the “floating ribs”) do not attach to the sternum at all. With increasing age, the sternal end of the rib becomes thinner and irregularly shaped compared to the smooth, rounded shape seen in young adults.

Appendicular Skeleton
Pectoral Girdle
The pectoral girdle consists of the clavicle and the scapula, and it serves as the proximal base of the upper limb as well as the anchor for the upper limb to the axial skeleton. The clavicle is an S-shaped bone, and it forms the strut that connects the scapula to the sternum (Figure A.31). The scapula is a large, flat bone with three angles (superior, inferior, and lateral) and three borders (medial, lateral, and superior). The lateral angle is noteworthy because it serves as the articulation for the head of the humerus of the upper limb at the glenoid cavity (or glenoid fossa; Figure A.32). The borders and the anterior and posterior surfaces of the scapula are sites of muscle attachment. The scapula also has three important projections for muscle and ligament attachments: the coracoid process anteriorly and superiorly; the acromion, which articulates with the lateral end of the clavicle; and the spine on the posterior aspect of the scapula.

Upper Limb
The bones of the upper limb skeleton include the humerus, radius, ulna, eight carpal (wrist) bones, five metacarpal (hand) bones, and 14 phalanges (finger bones). Each of these bones is described below along with several of the prominent features.
The humerus is the bone of the arm. On the proximal epiphysis of the humerus are attachment sites for muscles of the rotator cuff (greater tubercle and lesser tubercle). A major shoulder muscle (deltoid muscle) attaches to the humerus along the lateral aspect of the diaphysis at the deltoid tuberosity. On the distal epiphysis of the humerus, the medial epicondyle is an attachment site for muscles that flex the forearm, and the lateral epicondyle is an attachment site for muscles that extend the forearm (Figure A.33).

There are two bones of the forearm, attached to each other by a thick connective tissue interosseous membrane: the radius and the ulna (Figure A.34). The radius is lateral to the ulna in anatomical position (this is called supination of the forearm), but it crosses over the ulna when the wrist is rotated so that the thumb points medially (this is called pronation of the forearm). On the proximal end of the radius is the radial tuberosity, an attachment site for the biceps brachii muscle that will help supinate and flex the forearm; on the distal end of the radius is the styloid process, an attachment site for ligaments of the wrist. The ulna also has a styloid process, but unlike the one on the radius it does not have a relevant function. Instead, the important processes on the ulna are located proximally, and they include the olecranon process for the attachment of the triceps brachii muscle (a muscle that extends the forearm and arm) and the coronoid process for the attachment of the brachialis muscle (a muscle that flexes the forearm).

There are eight carpal bones that comprise the wrist, and they are organized into two rows: proximal and distal (Figure A.35). The proximal row of carpals (from lateral to medial) includes the scaphoid, lunate, triquetrum, and pisiform. The distal row (from lateral to medial) includes the trapezium, trapezoid, capitate, and hamate with its distinctive hamulus (hook) for muscle and ligament attachments. Distal to the carpal bones are the digital rays, each of which contains a metacarpal (hand) bone and three phalanges (proximal, middle, and distal) or finger bones. The exception to this rule is the thumb, which has fewer phalanges (proximal and distal, but no middle) than the other digits.

Pelvic Girdle
The pelvic girdle consists of the two os coxae and the sacrum that articulates with both, and it serves as the proximal base and anchor of the lower limb to the axial skeleton. Each os coxa comprises three bones that fuse together during growth: ilium, ischium, and pubis. These three bones fuse in a region called the acetabulum, which is the socket for the ball-and-socket hip joint (Figure A.36). The ilium, the flared superior portion of the pelvis, is the largest bone of the os coxa and serves as a major site of attachments for muscles from the abdomen, back, and lower limb. The ilium has several important features including the auricular surface, the surface where the ilium articulates with the sacrum. The auricular surface is used to estimate age at death as the surface progressively deteriorates with increasing age to appear coarse and porous. The greater sciatic notch is a large notch in the ilium that allows for several structures to leave the pelvis and enter the lower extremity, including the sciatic nerve. In females, the notch tends to be symmetrical whereas in males it tends to curve posteriorly (Nawrocki et al. 2018).

The ischium forms the posterior and inferior portion of the os coxa. There are two significant projections of note on the ischium: the ischial spine and tuberosity. The ischial spine is the attachment point for a major pelvic ligament and is located inferior to the greater sciatic notch of the ilium. The ischial tuberosity is the proximal attachment site for the hamstring muscles of the lower limb.
The anterior and medial portions of the os coxa are formed by the pubis. The pubis is a useful bone with which to sex a skeleton in a forensic context (Bass 2005; Buikstra and Ubelaker 1994). The body is the superior and medial portion of the pubis (Figure A.37). The body tends to be rectangular in cross-section in females and triangular in males. The bony projection that unites the ischium and pubis anteriorly is called the ischiopubic ramus. Females tend to display a thin and sharp ramus on the medial surface while the surface in males tends to be broad and blunt. The joint that unites the two pubic bones in the front of the pelvis is called the pubic symphysis, which is a structure commonly used in age estimation. In young adults, the surface is billowed, but it transitions to being smooth and porous with increasing age. The subpubic concavity is a depression inferior to the ischiopubic ramus. Females tend to exhibit a concavity while males tend to be straight. Finally, the large opening encircled by the pubis and ischium is called the obturator foramen. The shape of the foramen in females has been described as triangular while it is more likely to appear oval in males (Bass 2005).

Lower Limb
The bones of the lower limb skeleton include the femur, patella, tibia, fibula, seven tarsal (ankle) bones, five metatarsal (foot) bones, and 14 phalanges (toe bones). Each of these bones is described below along with several of the prominent features.
The femur is the bone of the thigh. On the proximal epiphysis of the femur are attachment sites for major hip and thigh muscles on the greater trochanter, lesser trochanter, and gluteal tuberosity (Figure A.38). The raised ridge on the posterior aspect of the femoral diaphysis is called the linea aspera, and it is a major attachment site for the quadriceps femoris muscles and other muscles, and it terminates distally by splitting into medial and lateral epicondyles, additional sites of muscle attachment. The distal epiphysis of the femur is marked by two rounded condyles that articulate with the proximal part of the tibia. The anterior surface of the distal femur articulates with the patella (kneecap), a bone that develops within the tendon of the quadriceps femoris muscle to enhance the function of the muscle. The patella does not articulate with the tibia.

There are two bones of the leg: tibia and fibula. The tibia is the robust, medial bone of the leg, and it is connected to the laterally positioned fibula by an interosseous membrane like in the forearm (Figure A.39). The proximal epiphysis of the tibia has two articular facets called tibial condyles that articulate with the femoral condyles. On the anterior surface of the proximal tibia is a raised projection called the tibial tuberosity, where the quadriceps muscle tendon attaches distally after containing the patella. On the distal epiphysis of the tibia is the medial malleolus, which articulates with the talus in the ankle joint. The lateral malleolus is a feature of the distal end of the fibula; the proximal end of the fibula articulates with the lateral portion of the proximal tibia.

There are seven tarsal bones that comprise the ankle (Figure A.40). The talus is the most superior of the tarsals, and it articulates with the distal tibia and distal fibula superiorly and with the calcaneus inferiorly. The calcaneus is the heel of the foot; it is the largest of the tarsals. On the posterior-most aspect of the calcaneus is the calcaneal tuberosity, which is the attachment site for the Achilles tendon of the posterior leg. Distal to the talus is the medially positioned navicular, the three cuneiform bones (medial, intermediate, and lateral), and the laterally positioned cuboid. Distal to the tarsals are the digital rays, each of which contains a metatarsal (foot) bone and three phalanges (proximal, middle, and distal) or toe bones. The exception to this rule is the big toe, which has fewer phalanges (proximal and distal, but no middle) than the other digits.

STATURE ESTIMATION FROM ADULT SKELETONS
In forensic contexts, it makes sense that scientists would want to estimate the height of the individual whose remains were recovered. It also is reasonable that bioarchaeologists would want to estimate stature (height), because body size is one of the most important variables in assessing physiological processes like heart rate and metabolic rate. Stature estimation equations have been developed in bioarchaeological and forensic contexts that rely on measuring length of bones like the cranium, vertebrae, long bones of the limbs, and so forth. These measurements are then input into these equations and stature can be estimated from the resulting solution (Auerbach and Ruff 2010; Lundy 1988).
DIFFERENCES BETWEEN ADULT AND SUBADULT SKELETONS
The adult skeleton consists of 206 bones. Each of these bones develops from a number of centers of ossification. It is estimated, then, that a baby is born with approximately 450 bones that grow from their centers of ossification and eventually become the 206 bones of the adult skeleton. For example, a typical long bone (e.g., tibia) has three centers of ossification: one primary center, the diaphysis; and two secondary centers, the epiphyses. In between epiphysis and diaphysis is an epiphyseal growth plate of cartilage that will remain unfused until postnatal growth is complete after puberty (and for some bones, well into adulthood). There is a relatively well documented order in which bones of the subadult postcranial skeleton reach full fusion, which is used as the basis for age estimation in forensic contexts (Bass 2005; White and Folkens 2000).
Similarly, the sutures between cranial bones in children are unfused, which allows skull growth to coincide with brain growth and provides a basis for age estimation based on suture fusion. The skulls of babies are marked by several fontanelles (soft spots), which are areas of the skull filled with membrane that has not been replaced with bone through intramembranous ossification.
Finally, the age of the subadult skeleton can be estimated based on teeth. All mammals develop two sets of teeth: deciduous (baby) teeth and permanent (adult) teeth; humans are no exception. Permanent human teeth were described immediately above. It is worth spending a few words to describe deciduous human teeth. At birth, humans usually display no teeth, but by about six months of age, the deciduous lower central incisors usually appear (see Bass 2005). When the complete sequence of deciduous teeth has erupted, there are five teeth in each quadrant: two incisors, one canine, and two molars. Deciduous incisors and canines are eventually replaced by their adult counterparts; deciduous molars are replaced by adult premolars, and there is no deciduous precursor to adult molars. The eruption patterns of deciduous and adult teeth is well documented and is used as in forensic contexts to estimate age (Bass 2005).
COMPARATIVE SKELETAL ANATOMY
Over the last six to seven million years, humans have been evolving to become more efficient at walking around on two limbs (bipedal locomotion), resulting in skeletal anatomy that is divergent from our closest living relative, the chimpanzee (Pan troglodytes). These differences can be seen in both the axial and appendicular skeletons. In the axial skeleton, for example, the foramen magnum in humans is more anteriorly positioned than that of chimpanzees, which places the vertebral column directly underneath the skull as opposed to behind it as it is in chimpanzees and other quadrupedal mammals. Chimpanzees also do not have an S-shaped curvature to their vertebral column; they simply retain the gentle primary kyphosis developed during the fetal period. Furthermore, they actually have one fewer lumbar vertebrae compared to humans, which results in a stiffer lower back.
As far as differences in the appendicular skeleton are concerned, the shape of the pelvic girdle is dramatically different in humans, where the ilium flares out laterally compared to the posterior flare of the ilium in chimpanzees. This reorganization of the pelvis has changed the function of two muscles, gluteus medius and gluteus minimus, from hip extensors in chimpanzees to hip abductors in humans. The angle of the femoral diaphysis is more oblique in humans because one of the demands of efficient bipedal locomotion is that humans require their knees to remain under their center of mass when they are standing on one limb during walking; in chimpanzees, the knees are not moved under their center of mass, so the femoral diaphysis is nearly vertical in orientation from the hip to the knee joint. In addition, humans have oversized hip and knee joints for their body size compared to chimpanzees, likely because they require more surface area to keep from damaging joint surfaces when they support their entire body mass on a single limb during walking. Chimpanzees spend more time engaging in climbing behavior than humans do, and they are known to have glenoid fossae and scapulae that are oriented more superiorly than humans, which allows them to support their body weight better on their upper limbs than humans can. Finally, the phalanges of the hand and foot are less anteroposteriorly curved in humans than they are in chimpanzees, and instead of having an opposable big toe like chimpanzees, humans have a big toe that is in line with the other digits and more efficient for bipedal locomotion.
Review Questions
- Which bony features of the pelvic girdle are relevant to estimating age and/or sex in forensic and bioarchaeological contexts? Give specific examples of how these features differ among sexes.
- What is the mechanistic difference between endochondral and intramembranous bone formation?
- Which bones articulate with the calcaneus? Which bones articulate with the humerus?
- Which elements of the skeleton belong to the axial skeleton versus the appendicular skeleton?
- Describe the axial and appendicular skeletal differences between humans and chimpanzees.
About the Authors
Jason M. Organ, Ph.D.
Indiana University School of Medicine

Jason M. Organ, Ph.D., is an assistant professor of Anatomy, cell biology, & physiology at the Indiana University School of Medicine and Co-editor and Writer for the Public Library of Science (PLOS) Science Communication Blog, where he advocates for the importance of storytelling and empathy in science communication. Dr. Organ earned his M.A. in anthropology from the University of Missouri and his Ph.D. in functional anatomy & evolution from Johns Hopkins University School of Medicine. He has published over 30 peer-reviewed research papers on evolutionary and mechanical adaptations of bone and muscle in scientific journals and over 40 peer-reviewed teaching modules in digital human anatomy references. Dr. Organ recently completed a three-year term as an elected member of the Board of Directors of the American Association for Anatomy (AAA), where he advocated for the importance of effective science communication and public outreach at the association level, with an emphasis on connecting with policy makers to ensure sufficient federal funding for science. In 2018, Jason received the prestigious AAA Basmajian Award for excellence in teaching gross anatomy and outstanding accomplishments in biomedical research and scholarship in education. Follow Dr. Organ on Twitter: @OrganJM
Jessica N. Byram, Ph.D.
Indiana University School of Medicine

Jessica N. Byram, Ph.D., is an assistant professor of Anatomy, cell biology, & physiology at the Indiana University School of Medicine (IUSM). Jessica earned her M.S. in human biology with a focus in forensic anthropology from the University of Indianapolis and her Ph.D. in anatomy education at IUSM. Jessica is the director of the anatomy education track Ph.D. program at IUSM. Her research interests include medical professionalism, investigating professional identity formation in medical students and residents, and exploring how to improve the learning environments at medical institutions.
References
Auerbach, Benjamin M., and Christopher B. Ruff. 2010. “Stature Estimation Formulae for Indigenous North American Populations.” American Journal of Physical Anthropology 141 (2): 190–207.
Bass, William. 2005. Human Osteology: A Laboratory and Field Method. Springfield, IL: Charles C. Thomas.
Buikstra, Jane E., and Douglas H. Ubelaker. 1994. Standards for Data Collection From Human Skeletal Remains. Arkansas Archaeological Survey Research Series, 44. Fayetteville, AR: Arkansas Archeological Survey.
Burr, David B., and Jason M. Organ. 2017. “Postcranial Skeletal Development and Its Evolutionary Implications.” In Building Bones: Bone Formation and Development in Anthropology, edited by Christopher J. Percival and Joan T. Richtsmeier, 148–174. Cambridge, UK: Cambridge University Press.
Federative Committee on Anatomical Terminology (FCAT). 1998. Terminologia Anatomica: International Anatomical Terminology. Stuttgart, Germany: Georg Thieme Verlag.
Lundy, J. K. 1988. “A Report on the Use of Fully’s Anatomical Method to Estimate Stature in Military Skeletal Remains.” Journal of Forensic Sciences 33 (2): 534–539.
Nawrocki, Stephen P., Krista E. Latham, Thomas Gore, Rachel M. Hoffman, Jessica N. Byram, and Justin Maiers. 2018. “Using Elliptical Fourier Analysis to Interpret Complex Morphological Features in Global Populations.” In New Perspectives in Forensic Human Skeletal Identification, edited by Krista E. Latham, Eric J. Bartelink, and Michael Finnegan, 301–312. London: Elsevier/Academic Press.
Organ, Jason M. 2017. “Vertebral Column.” Amirsys Anatomy Reference Center. Salt Lake City: Elsevier. https://app.anatomyreferencecenter.com/
White, Tim D., and Pieter A. Folkens. 2000. Human Osteology, 2nd Edition. New York: Academic Press.
———. 2005. The Human Bone Manual. London: Elsevier.
Figure Attributions
Figure A.1 Regions of the Human Body (Anatomy & Physiology, Figure 1.12) by OpenStax has been modified (labels removed) and is used under a CC BY 4.0 License.
Figure A.2 Planes of the Body (Anatomy & Physiology, Figure 1.14) by OpenStax has been modified (some labels modified) and is used under a CC BY 4.0 License.
Figure A.3 Directional Terms Applied to the Human Body (Anatomy & Physiology, Figure 1.13) by OpenStax is used under a CC BY 4.0 License.
Figure A.4 Anatomy of a Long Bone (Anatomy & Physiology, Figure 6.7) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.5 Bone Cells (Anatomy & Physiology, Figure 6.11) by OpenStax is used under a CC BY 4.0 License.
Figure A.6 Classifications of Bones (Anatomy & Physiology, Figure 6.6) by OpenStax is used under a CC BY 4.0 License.
Figure A.7 Intramembranous Ossification (Anatomy & Physiology, Figure 6.16) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.8 Endochondral Ossification (Anatomy & Physiology, Figure 6.17) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.9 Pathways in Calcium Homeostasis (Anatomy & Physiology, Figure 6.24) by OpenStax is used under a CC BY 4.0 License.
Figure A.10 Axial and Appendicular Skeleton (Anatomy & Physiology, Figure 7.2) by OpenStax is used under a CC BY 4.0 License.
Figure A.11 Parts of the Skull (Anatomy & Physiology, Figure 7.3) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.12 Anterior View of Skull (Anatomy & Physiology, Figure 7.4) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.13 Lateral View of Skull (Anatomy & Physiology, Figure 7.5) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.14 Temporal Bone (Anatomy & Physiology, Figure 7.7) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.15 External and Internal Views of Base of Skull (Anatomy & Physiology, Figure 7.8) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.16 Sphenoid Bone (Anatomy & Physiology, Figure 7.10) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.17 Ethmoid Bone (Anatomy & Physiology, Figure 7.12) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.18 Maxillary Bone (Anatomy & Physiology, Figure 7.14) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.19 Bones of the Orbit (Anatomy & Physiology, Figure 7.16) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.20 Nasal Septum (Anatomy & Physiology, Figure 7.17) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.21 Lateral Wall of Nasal Cavity (Anatomy & Physiology, Figure 7.13) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.22 Hyoid Bone (Anatomy & Physiology, Figure 7.19) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.23 Isolated Mandible (Anatomy & Physiology, Figure 7.15) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.24 Parts of a Typical Vertebra (Anatomy & Physiology, Figure 7.23) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.25 Cervical Vertebra (Anatomy & Physiology, Figure 7.25) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.26 Rib Articulation in Thoracic Vertebrae (Anatomy & Physiology, Figure 7.27) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.27 Lumbar Vertebrae (Anatomy & Physiology, Figure 7.28) by OpenStax is used under a CC BY 4.0 License.
Figure A.28 Sacrum and Coccyx (Anatomy & Physiology, Figure 7.29) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.29 Vertebral Column (Anatomy & Physiology, Figure 7.20) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.30 Thoracic Cage (Anatomy & Physiology, Figure 7.32) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.31 Pectoral Girdle (Anatomy & Physiology, Figure 8.3) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.32 Scapula (Anatomy & Physiology, Figure 8.4) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.33 Humerus and Elbow Joint (Anatomy & Physiology, Figure 8.5) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.34 Ulna and Radius (Anatomy & Physiology, Figure 8.6) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.35 Bones of the Wrist and Hand (Anatomy & Physiology, Figure 8.7) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.36 Pelvis (Anatomy & Physiology, Figure 8.12) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.37 The Hip Bone (Anatomy & Physiology, Figure 8.13) by OpenStax has been modified (some labels removed) and is used under a CC BY 4.0 License.
Figure A.38 Femur and Patella (Anatomy & Physiology, Figure 8.16) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
Figure A.39 Tibia and Fibula (Anatomy & Physiology, Figure 8.18) by OpenStax has been modified (one label removed) and is used under a CC BY 4.0 License.
Figure A.40 Bones of the Foot (Anatomy & Physiology, Figure 8.19) by OpenStax has been modified (some labels modified or removed) and is used under a CC BY 4.0 License.
